





- Microbial Amelioration of Acid Mine Drainage Impaired Soil using the Bacterial Consortia of Klebsiella sp. and Raoultella sp.
Seon Yeong Park1·Gi Won Lee2·Chang Gyun Kim1,2,*
1Program in Environmental and Polymer Engineering, INHA University, Incheon 22212, Korea
2Department of Environmental Engineering, INHA University, Incheon 22212, KoreaThis article is an open access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Acid mine drainage (AMD) resulting from pyrite oxidation in mining areas, subsequently leads to soil acidification accompanied by lowering pH and high concentration of metals and metalloids in its surrounding environment. Regarding to this, the microbial amelioration has been considered as a promising option for a more cost-effective and eco-friendlier countermeasure, compared to the use of alkaline chemicals. This study was aimed to evaluate influencing factors in microbially-mediated amelioration of acidic soil spiked by simulated AMD. For this, microcosm experiments were conducted by acid-neutralizing bacterial consortium (dominated by Klebsiella sp. and Raoultella sp.) under the various conditions of AMD spikes (0-2,500 mg SO42-/L), together with acidic mine soil (0-100 g) or sphagnum peat (0-5 g) in the 200 mL of nutrient medium. The employed bacterial consortium, capable of resisting to high level of sulfate concentration (up to 1,500 mg SO42-/L) in low pH, generated the ammonium while concomitantly reduced the sulfate, subsequently contributing to the effective soil stabilization with an evolution of soil pH up to neutral. Furthermore, it demonstrates that suitable condition has to be tuned for successful microbial metabolism to facilitate with neutralization during practical application.
Keywords: Acid mine drainage, Acid-neutralization, Bioremediation, Deamination, Sulfate
Acid mine drainage (AMD) originated from either active or abandoned mining sites is characterized by low pH (< 4) and high concentration of potentially toxic dissolved metals, metalloids and sulfate, provoking severe pollution (Kefeni et al., 2017; Zhang et al., 2019) in terms of soil acidification and following destruction of the ecosystem in the surrounding areas. In particular, while passing through neighboring rock layers, AMD specifically elevates the level of concentration of dissolved metals and metalloids such as zinc (Zn), iron (Fe), manganese (Mn), cobalt (Co), cadmium (Cd), nickel (Ni), aluminum (Al), and sulfate ion (SO42-) (Asta et al., 2010; Luptakova et al., 2012), leading to the contamination of surface water and natural groundwater (Fávere et al., 2004; Núñez-Gómez et al., 2018). Furthermore, the combination of high acidity and toxic materials such as Al and Mn, adversely affect plant and microorganism growth. For instance, in acidic soil, Al is solubilized to its trivalent cation, Al3+, which limits the availability of indispensable soil nutrients (e.g., phosphorous (P), molybdenum (Mo), and magnesium (Mg)) to be taken for plant growth, along with exacerbating the continuous loss of basic cations such as calcium (Ca2+) and potassium (K+) (Riaz et al., 2018). It was reported that root elongation can be inhibited even at micromolar concentrations of Al3+ because of which can in turn impair uptake of water and nutrients into it, thus contributing to poor growth and productivity of crops (Ma and Furukawa, 2003). Similarly, Mn is also considered as the major limiting factor to hindering crop cultivation in acidic soil because continuous Mn accumulation in plant obstructs essential nutrient uptake, eventually inducing vegetation to be prohibited for efficient photosynthesis rate (Huang et al., 2016).
Alkaline chemicals such as limestone (CaCO3) and hydrated lime (Ca(OH)2) have been used to effectively remove the various metals (e.g., Al, As, Cd, Co, Cu, Fe, Mn, Ni, and Zn) and sulfate contained in the AMD (Olds et al., 2013; Tolonen et al., 2014; Vadapalli et al., 2015), and improve soil fertility if properly used (Goulding, 2016). Nonetheless, its implementation has been quite slow because of the economic challenges involved with continuous supply of alkaline chemicals to arable land (Haynes and Mokolobate, 2001) or AMD treatment process (RoyChowdhury et al., 2015). Furthermore, chemically inclined AMD treatment processes annually produce huge quantities of waste sludge while consuming the operating and maintenance costs (Kefeni et al., 2017). Moreover, incessant liming can often induce soil nutrient imbalance, which can change soil microbial diversity and their activity (Marschner et al., 2003).
On the contrary, biological neutralization approach has been investigated as a more economical and eco-safe process compared to conventional methods (Ogbughalu et al., 2017; RoyChowdhury et al., 2015). Bioaugmentation of iron oxidizing bacteria (Sharma et al., 2020), iron and sulfate-reducing bacteria (SRB) (Gupta and Sar, 2020; Sánchez-Andrea et al., 2014) or adsorption of microbial fuel cells (Leiva et al., 2016; Tang et al., 2016) have been proposed as the attractive strategies to treat AMD. Moreover, the microbial decarboxylation and deamination of organic matters can increase soil pH by consuming protons and generating ammonium ion (NH4+), respectively (Okai et al., 2017; Yan and Schubert, 1996; Yuan et al., 2011). For instance, indigenous soil microbial isolates can be tolerant of high acidity, increasing the pH from 4 to 6.86, while concurrently increasing the ammonium nitrogen concentration up to 367.5 mg NH4+-N/kg of the soil slurry within 6 days (Park et al., 2016). SRB convert nitrogen compounds (e.g., explosives) to amino acids, and subsequently deaminate them to produce ammonia (Boopathy et al., 1998), which can create an alkaline environment around the cell as producing NH4+ and OH- ions from the binding with water in the strict anaerobic condition (Rodriguez-Navarro et al., 2003). In the meantime, NH4+ could be also produced through the denitrification of nitrate by nitrate-reducing and sulfide-oxidizing bacteria (Hubert and Voordouw, 2007). Nevertheless, the determination of their acid neutralization capacity has been still challenged owing to the various environmental factors (including numerous heterogenous soil properties and type of soil acid precursors). For this reason, the remediation technology of soil impaired by AMD containing high SO42- concentration has been less developed than that of the AMD itself (Qin et al., 2019).
This study, therefore, aimed to evaluate the influencing factors on microbially-mediated amelioration of AMD-impaired acidic soil using the heterogenous acid-neutrali- zing bacterial consortium which was isolated from general soil. For this purpose, the experiment carried out under the various conditions, in terms of different AMD spikes (0-2,500 mg SO42-/L), or proportion of mimicked acidic mine soil (0-100 g) and sphagnum peat in soil (0-5 g) while in the constant volume of nutrient medium kept, for which a temporal variation of pH, NH4+ production yield, and microbial enzymatic activity (i.e., dehydrogenase activity) have been monitored. Ultimately, possible influencing factors were statistically analyzed to determine the effect of them on neutralizing acidic pH of soil originated from AMD.
2.1. Isolation of acid-neutralizing microbial consortium and its identification
Soil samples collected from a 5 cm depth below ground surface in garden site of a metropolitan area (37°45' 07.92"N, 126°65'59.23"E, Michuhol-gu, Incheon, Korea) were sieved using a 10-mesh stainless steel sieve, and then their physicochemical properties were analyzed according to each of standard analytical protocols. Briefly, pH was measured by pH meter (BP 3001, Trans Instruments, Singapore) after mixing 5 g of dried soil into the 25 mL of deionized water, while their moisture and organic content were determined by comparing the weight of them with those of dried at 105oC or ignited at 550oC, respectively. In the meantime, 5 g of soil samples were suspended in the 100 mL of autoclaved Luria-Bertani (LB) broth (containing 10 g/L of BactoTM Tryptone, 5 g/L of BactoTM Yeast Extract, and 8 g/L of NaCl; adjusted to pH 7.0) after which the microbes were cultivated for 24 hours at 30oC in an orbital shaking incubator (VS-8480SF, Vision Scientific Co. Ltd., Korea) with a shaking rate of 160 rpm. After allowing soil to settle down for 30 minutes, 10 mL of supernatant was transferred into 100 mL of acidic LB broth previously adjusted to pH 4.0 by diluted HCl solution and repeatedly cultivated in the same incubation condition. Concomitantly, their acid-neutralizing capacities were simply verified by streaking them onto the acidic LB agar medium containing 0.01% (w/v) of methyl red indicator which helps to distinguish via the color change of medium from pink to yellow depending on the pH evolution. After incubating them at 30oC in a static incubator (VS-8480, Vision Scientific Co. Ltd., Korea), colonies formed on the color-changed zone were individually transferred into LB broth and enriched. Each bacterial strain was harvested by centrifuging at 10,000 × g for 5 minutes (Centrifuge 5415D, Eppendorf, Germany).
For the bacterial identification, their genomic DNA were extracted using a Soil Fast DNATM Spin Kit (MP Bio- medicals, USA) according to the manufacturer’s instruction. They were immediately amplified based on the con- ventional polymerase chain reaction (PCR) using BioFACTTM Taq DNA polymerase kit (BioFACT, Korea) with a universal primer set of 27F (5'-AGA GTT TGA TCM TGG CTC AG-3') and 1492R (5'-TAC GGY TAC CTT GTT ACG ACT T-3', synthesized by BIONEER, Korea). PCR was programmed with the initial denaturation (95oC, 5 minutes), 35 cycles of denaturation (95oC, 1 minutes), annealing (54oC, 1 minutes), extension (72oC, 1 minutes), and further holding at 72oC for 10 minutes in a thermal cycler (FTGENE2D, Techne, UK). The resulting amplicons were purified using a HiGeneTM Gel & PCR Purification System (BioFACT, Korea) and they were confirmed by electrophoresis in 1% (w/v) agarose (for molecular biology, SIGMA) gel in a Tris-acetate-EDTA buffer (composed of 40 mM Tris-acetate and 1 mM EDTA). The 16S rRNA genes were sequenced in the ABI Prism® 3100 Genetic Analyzer (Hitachi, Japan), after which they were inde- pendently compared to match with the 16S rRNA sequence database in the National Center for Biotechnology Infor- mation (NCBI) through the Basic Local Alignment Search Tool (BLAST) (Ahn and Kim, 2015).
In addition, prior to the microbial amelioration test, the minimum pH where microbes actively metabolize for pH evolution was determined by inoculating the bacterial culture onto the LB agar plates previously acidified in various acidic pH ranges (i.e., pH 3, 4, 5, and 6) using AMD stock solution.
2.2. Evaluation for inhibitory effect of AMD on microbial activity
The effects of AMD on microbial growth and following acid neutralization were tested by increasing initial doses of simulated AMD, which are made up by mixing equal volume of 2 M ferrous sulfate (FeSO4.7H2O, > 98.0%, Duchefa Biochemie, Netherlands) solution and 2 M sulfuric acid (95.0%, Samchun Chemicals, Korea) solution. It was prepared in the same manner as addressed by the pyrite oxidation mechanism: 2FeS2 + 7O2 + 2H2O {{{EQUATIONS}}} 2Fe2+ + 4SO42- + 4H+ (Costa and Duarte, 2005). For that, AMD were added to the final concentrations of 0, 500, 1,000, 1,500, 2,000, and 2,500 mg SO42-/L into the respective six Erlenmeyer flasks containing 200 mL of LB medium, and then further adjusted their pH to 4.0 ± 0.5 using HCl solution or pulverized calcium carbonate (CaCO3, Duchefa Biochemie, Netherlands). The acid-neutralizing microbial consortium was inoculated when its optical density at the wavelength of 600 nm (OD600) was reached up to 1.68, and then they were incubated for 7 days at 30oC in the dark shaking incubator with an agitation rate at 160 rpm. The test was carried out in duplicate for which pH, NH4+ concentration, and microbial activity as of OD600 were daily monitored. In addition, the aliquot was acidified to pH 2.0 by nitric acid (HNO3, 60.0%, Samchun Chemicals, Korea) to remove the residual organic matters such as cellular debris or metabolites, and then its SO42- concentration was quantified using ion chromatography (ICS-3000, Dionex, USA).
2.3. Microbial amelioration test of AMD-impaired soil
The microbial amelioration was tested by the effect of varying dose of AMD-impaired soil together with different mass fraction of sphagnum peat. First, the effect of dose of AMD-impaired soil was evaluated in the constant volume of nutrient medium amended with the acid-neutralizing bacterial consortium. Herein, the soil was synthesized by blending industrial sand (SiO2, particle size range of 50-200 μm, Kyungin Material, Korea), kaolinite clay (H2Al2Si2O8. H2O, Samchun Chemicals, Korea), and sphagnum peat with a proportion of 70, 20, and 10% (w/w), respectively, as suggested in Organization for Economic Cooperation and Development (OECD) Guideline for Testing of Chemicals No. 207 (OECD, 1984), in order to neglect the environ- mental variables derived from field soil. These materials were individually autoclaved at 120oC for 15 minutes (MLS-3780, Sanyo, Japan), and then they were intermixed thoroughly with a moisturization at least 48 hours before the application to stabilize their acidity. And thereafter, 200 mL of LB broth was infused into each Erlenmeyer flasks containing different amounts of synthesized soils (0, 1, 10, 25, 50, 60, 75, and 100 g) and then pH was adjusted to pH 4.0 ± 0.5 using AMD stock solution. For that, AMD was added with different concentrations of 787.2, 777.6, 672, 480, 288, 192, 96, and 48 mg SO42-/L into soil slurry containing 0, 1, 10, 25, 50, 60, 75, and 100 g of soil, respectively. Second test was followed with varying mass of sphagnum peat (i.e., 0, 0.5, 1.0, 2.5 and 5.0 g) while maintaining 7 g of industrial sand and 2 g of kaolinite clay in the 200 mL of LB broth, respectively. At this time, since the sphagnum peat can also decrease the soil pH (Lee et al., 2015), the final pH of medium was adjusted to pH 4.0 ± 0.5 using pulverized CaCO3 while maintaining the AMD concentration to be the same of 787 mg SO42-/L. For all bioaugmented sets, the exponential-phase microbial culture (OD600 = 1.68) was at the same way inoculated and incubated at 30oC with an agitation rate of 160 rpm in the shaking incubator (VS-8480SF, Vision Scientific, Korea). During the amelioration period of 6 days, the effects of each influencing factors on microbial amelioration were com- pared by monitoring the variation in pH, NH4+ production yield and dehydrogenase activity. Simultaneously, abiotic control sample was set up with the same manner, except for bacterial inoculation.
2.4. Analytical procedures: pH, NH4+ concentration and microbial activity
During the microcosm test, the samples were taken from each reactor to determine the pH, NH4+ production yield and microbial metabolic activity. First, pH was measured using a pH meter (JENWAY 3510, JENWAY, UK) and NH4+ concentration was determined by the Nessler method with a supernatant taken after centrifugal separation of them at 3,000 × g for 15 minutes (HA-1000-3, Hanil Science Co. Ltd., Korea). Briefly, 50 mL of diluted supernatant was added with 1 mL of Nessler reagent, 3 drops of mineral stabilizer, and 3 drops of polyvinyl alcohol dispersing agent (Hach, USA) after which its absorbance was then measured at 425 nm using an UV-Vis spectrometer (UV-3300, Humas Co. Ltd., Korea) within 30 seconds. In respect to the metabolic activity, OD600 was simply measured using a UV-Vis spectrometer at the wavelength of 600 nm, while the dehydrogenase activity was determined through the in vivo oxidoreduction of 2-(4-Iodophenyl)-3-(4-nitrophenyl)-5-phenyltetrazolium chloride (INT, C19H13ClIN5O2, TCI, Japan). For modified INT assay (Trevors, 1984), 500 μL of sample was centrifuged at 10,000 × g for 5 minutes, and the harvested cell pellet was agitated with 500 μL of 0.2% (w/v) INT dissolved in 200 mM phosphate buffer (pH 7.6) solution and 100 μL of 1% (w/v) sucrose (C12H22O11, Daejung Chemicals, Korea) solution. Incubating them in a dark incubator at 30oC for 30 minutes, 50 μL of concentrated HCl (35.0~37.0%, Samchun Chemicals, Korea) and 1 mL of 1,4-dioxane (anhydrous, 99.8%, Sigma, USA) were further added to extract red INT formazan, and then its absorbance was immediately measured at the wavelength of 481 nm using UV-Vis spectrometer. The obtained absorbance was converted into the degree of dehydrogenase activity referring to the concentration of iodonitrotetrazolium violet formazan (INF) given here. All analytical procedures were promptly conducted after taking samples to avoid the possible distorted interpretation.
2.5. Statistical analyses
In this study, the experimental data were expressed with the mean value and standard deviation, in which they were descriptively plotted using Microsoft Excel to compare the varying conditions. Additional statistical analyses were carried out using R version 3.6.3 (The R Foundation, Austria). In that, the correlation matrix was built up for the interaction among the microbial metabolic activity, pH evolution and NH4+ production, with Pearson’s correlation coefficients. Moreover, two-way analysis of variance (ANOVA) was employed to assess the effect of soil contents and incubation times on microbial amelioration leading to pH evolution. In addition, Tukey’s HSD Post-hoc comparison test was adopted to validate the significant difference in the mean value among the treatment groups. In the long run, the significance level was set up at p < 0.05 for all statistical analyses.
3.1. Identification of acid-neutralizing microbial con- sortium
Soil sample collected from the garden site was analyzed for pH 7.6, of which moisture and organic content were 5.6% and 4.1%, respectively. From this, heterotrophic acid-neutralizing bacterial strains were individually isolated to use as a biological soil ameliorant because there are very broad bands of unknown reasons to be involved in acidification of natural soils including AMD impaired soil. Their types of species were identified based on 16S rRNA gene sequencing technique referring from ascertaining their taxonomic positions. Potential phylogenetic matches were sorted out with at least 98% of similarity in comparing to the query-subject alignments in nucleotide database sequences with accession numbers from NCBI-BLAST. From this, they mainly belonged to five species of Citrobacter freundii ATCC 8090 (NR028894), Enterococcus hirae ATCC 9790 (NR075022), Klebsiella oxytoca ATCC 13182 (NR118853), Kluyvera ascorbata ATCC 33433 (NR028677), and Raoultella ornithinolytica ATCC 31898 (NR114502), with which their relative abundance was 6.3, 10.9, 42.6, 5.8 and 34.4%, respectively, after scrutinizing the colonies formed on the agar plates. Among them, Citrobacter sp. can reduce sulfate to sulfide (Qiu et al., 2009), or nitrate and nitrite to ammonia (Rehr and Klemme, 1989) in oxidizing organic compounds such as formaldehyde and formate under strict anaerobic condition consequently contributing to both AMD-induced metal precipitation (Liu et al., 2018; Qiu et al., 2009) and pH increment by generating alkalinity (Liu et al., 2018). Qiu et al. (2009) demonstrated that Citrobacter freundii strain DBM are flexible to switching its metabolism depending on the presence or the absence of oxygen, from which they can in turn increase pH by releasing carbon dioxide (CO2) under their aerobic respiration. Meanwhile, Raoultella spp. produces ammonia via in vivo urease reaction, which can resist acidic environment (Sugimori et al., 2013), and subsequently leads to increasing soil pH to neutral. Klebsiella sp. has been widely adopted to either degrade various organic pollutants such as chlorpyrifos (Sasikala et al., 2012) and nitrobenzene (Wang et al., 2012), or produce bioethanol through co-fermentation of cellobio- nate and glycerol (Tao et al., 2019). As well as, the microbe contributes to the remediation of Mn-laden wastewater through oxidization and insolubilization of bivalent Mn ions (Mn2+) under the mid-alkaline condition, with a slight increase in pH (Barboza et al., 2018).
In the meantime, prior to the microbial amelioration test of AMD-impaired soil, we determined the minimum pH in which the given acid-neutralizing bacterial consortium can be enough to maintain their metabolic activities. Being exposed in acidic LB agar plates adjusted to pH 5 and 6 using AMD stock solution, they made a rapid increase in pH up to neutral, while at the initial pH 4, there was an acclimation period of one day needed, and thereafter the pH has been occurred to continuously increase for the following 3 days. However, at lowest initial pH 3, there was no observation of bacterial colonies and color change of medium. Thus, the bacterial consortium can be agreeably used as the soil ameliorant to neutralize the acidic soil impacted by AMD around pH 4, where the metabolic reaction of given bacterial strains was initiated.
3.2. Inhibitory effect of AMD on microbial growth
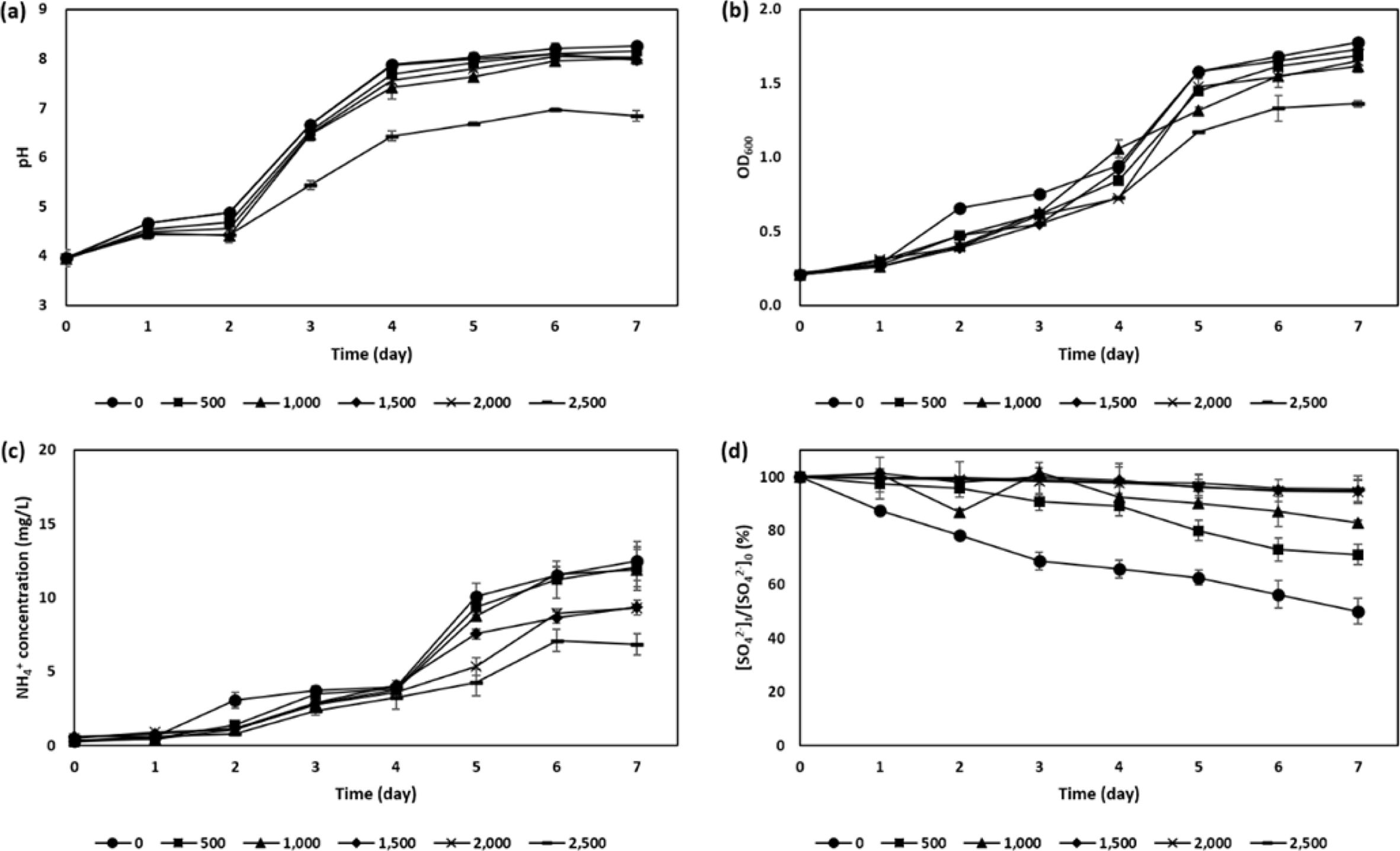
The minimum inhibitory concentration of AMD for a given acid-neutralizing bacterial consortium was determined by daily monitoring pH, microbial optical density (OD600), the concentration of ammonium (NH4+) and sulfate ion (SO42-) during the incubation time of period at the varying levels of AMD concentration of 0, 500, 1,000, 1,500, 2,000 and 2,500 mg SO42-/L, respectively (Fig. 1). As a result, pH values have been proportionally increased upon the increment of OD600 (Fig. 1 (a) and (b)), on the condition that the AMD concentration was kept up less than 2,000 mg SO42-/L. On the contrary, AMD concentration exceeding 2,000 mg SO42-/L overall repressed the microbial growth rates, which can in turn dwindle following neutralization capacity in it. In other words, at the termination of incubation period of 7 days, microbial growth and pH were reduced by 24.0% and 18.1%, respectively, at 2,500 mg SO42-/L, comparing to those without AMD, while the mean decrease rate for them was 2.4% and 5.3% in the lower AMD concentration, respectively. Meanwhile, NH4+ pro- duction yield was increased up to 13.02 ± 0.24 mg/L, 10.27 ± 0.47 mg/L and 6.95 ± 0.00 mg/L within the three different AMD concentration ranges in terms of relatively lower (0-1,000 mg SO42-/L), medium (1,500 and 2,000 mg SO42-/L), and higher (2,500 mg SO42-/L) levels, respectively, as presented in Fig. 1 (c). Furthermore, as the variation of SO42- concentration versus initial doses (Fig. 1 (d)) was delineated that the maximum reduction rate was 32.4% at the initial dose of 500 mg SO42-/L, followed by 18.9% at that of 1,000 mg SO42-/L. However, in the absence of AMD, the slope of sulfate reduction was suggested to be the steepest comparing to other experimental conditions adding up AMD. Such a greater extent of difference in the initial versus the followed temporally decreasing sulfate concentrations might be originated from an artifact contained as in one of the medium. Herein, a smaller quantity of SO42-, approximately 30 mg SO42-/L, might be coming from LB medium composites, tryptone and yeast extract. It has been confirmed using FTIR analysis that they have functional groups containing sulfuric compounds such as -CS-NH, >C=S, >SO and SO2N detected at the wavenumber of 1457 cm-1 and 1080 cm-1 in their spectra. On the contrary, the sulfate reduction rate was rapidly decreased to be only 5-7% at AMD concentrations more than 1,500 mg SO42-/L, implying that the microbial metabolism relevant to the sulfate ion can be inhibited by high dose of sulfate, even though the microbes still have been active to increase pH.
3.3. Microbial amelioration of soil impacted by AMD
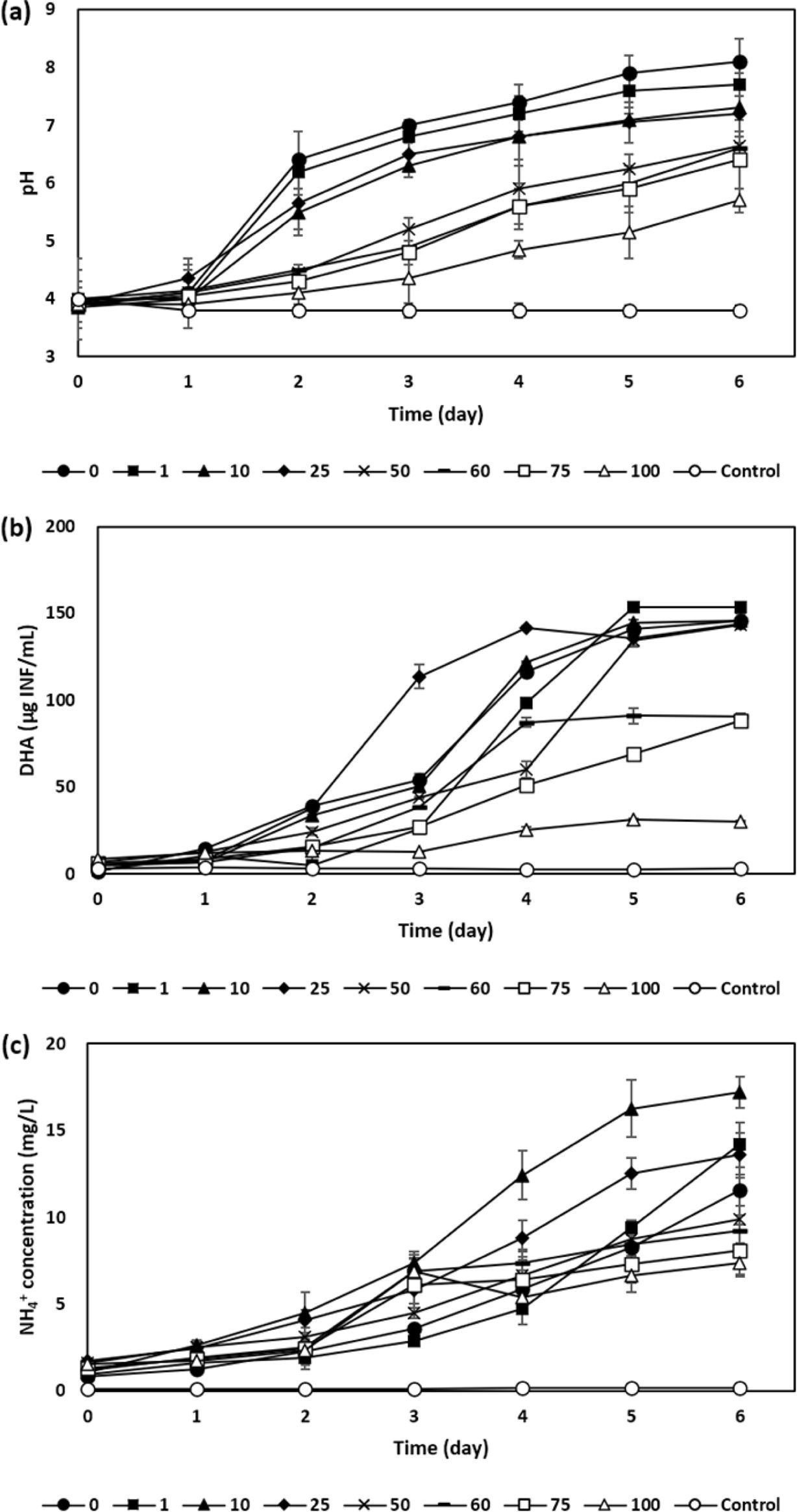
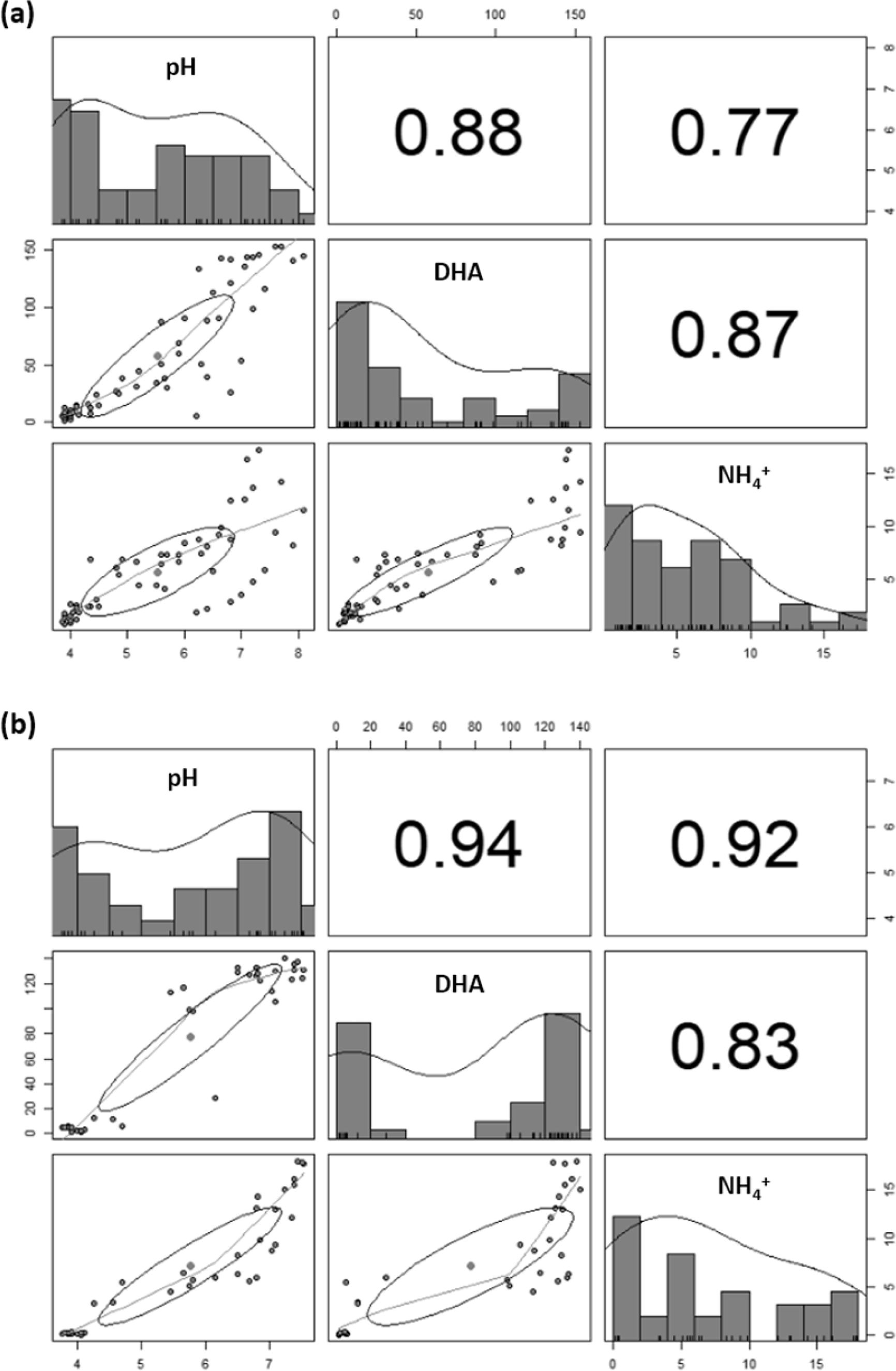
In this study, the amount of added acid mine soil and its fraction of sphagnum peat were investigated on the microbial metabolic activities and neutralization of soil slurry. First experiment was conducted on a varying amounts of acid mine soil slurries of which the solution pH was adjusted to 4.0 by spiking AMD. In this regard, dose of AMD was inversely proportional to the amount of synthesized soil with a high correlation coefficient of -0.98 with a significant difference (p< 0.05). From them, the final pH increased up to a wide range from 5.7 ± 0.2 (for 100 g of acid mine soil slurry) to 7.7 ± 0.2 (for 1 g of acid mine soil slurry) after 6 days of test period (Fig. 2 (a)), which was supposed to be induced by a microbial metabolism in terms of dehydrogenase activity (Fig. 2 (b)). Comparing to this, the absence of soil particles had increased up to the highest value of pH 8.10 ± 0.4. In the meantime, NH4+ concentration for 0 g of soil mass increased from 0.84 to 11.54 mg/L for 6 days of incubation, while the highest concentration of 17.22 mg/L was observed to be for 10 g/L of soil added at 6th day (Fig. 2 (c)). This difference between temporal increasing value of pH and produced NH4+ concentration can be attributed to enhancing self-protective microbial activity against these acidic conditions, which in turn improves NH4+ productivity (Nancharaiah et al., 2017; Wang et al., 2012) by fortifying their microbial granules shrouded around the soil particles. The correlation analysis among these parameters (Fig. 4 (a) also delineated that the relation between the microbial enzymatic activity (as of DHA) and NH4+ production yield has a higher correlation efficient (R) of 0.87 than that of relation between NH4+ production and pH evolution (R=0.77). Nonetheless, the incremental addition of synthesized soil into the liquid medium exceeding 50 g/L rather decreased the NH4+ production rate. It was seen that the competition between sphagnum peat and kaolinite against pH variable cationic of ammonium ion, which can be very dependently influenced by either the release of the proton (H+) or hydroxyl ion (OH-) according to varying pH condition (Chemeda et al., 2018). Thereafter, they can eventually inhibit microbial growth and metabolic activity, leading to the reduction of production rate of NH4+ relevant to agreeable neutralization (Freeman and Lock, 1992). In addition, sphagnum peat is mainly composed of humic substance such as humic acid, fulvic acid, and humin that have diverse functional groups such as carboxylic, alcoholic, phenolic, and hydroxylic groups (Smilek et al., 2015). These functional groups strongly dependent on pH; at higher alkaline pH, they are highly related to metal adsorption (Qi et al., 2017; Smilek et al., 2015) or the proton attraction (Yan and Schubert, 1996), while at lower pH, they are conversely protonated (Alice et al., 2016).
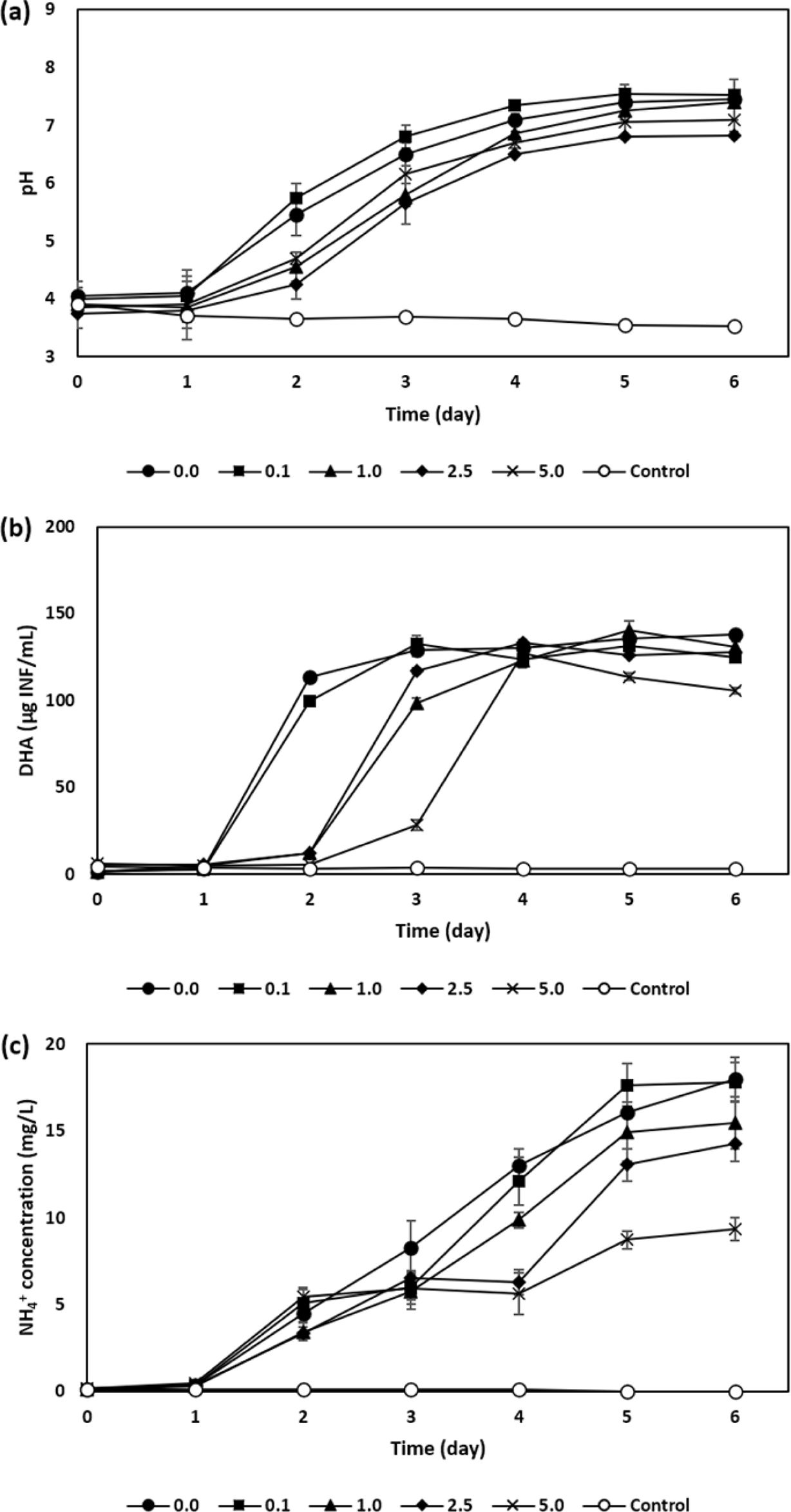
In line with this, to determine the effect of sphagnum peat on the given biological acid neutralization, varying amounts of sphagnum peat were added into a medium containing 7 g of industrial sand and 2 g of kaolinite, respectively, showing highest improvement in increasing pH compared to that without those of treatment as for the former experimental set-up. From that, pH in all bioaugmented sets had been increased up to in a range from pH 6.82 ± 0.0 to 7.52 ± 0.2 after 6 days of incubation (Fig. 3 (a)). Higher pH increase rate was obtained when 0.1 g of sphagnum peat was added, consequently generating up to approximately 17.63 mg/L NH4+ (Fig. 3 (c)). Meanwhile, the microbial activity and pH evolution were commonly suppressed when the sphagnum peat was more increasingly added due to the inhibitory effect of humic substances being engaged (Tejeda-agredano et al., 2014; Tikhonov et al., 2010). In the meantime, there was a higher correlation coefficient obtained at 0.83-0.94 as shown in Fig. 4 (b).
In addition, the two-way analysis of variance (ANOVA, Table 1) test revealed that there was a significant effect of amount of acidic soil and incubation times on amenable pH evolution (p < 0.001). It was also shown that sphagnum peat could less effect on pH variation with an increasing incubation time in the given experimental condition where 7 g of industrial sand and 2 g of kaolinite were added. Along with, the Turkey’s HSD Post-hoc comparison test also demonstrated that pH evolutions in all microcosm tests contained acidic mine soil slurry were quite different from those without microbes as of the control.
|
Fig. 1 Temporal variations in (a) pH, (b) microbial density as of OD600, (c) NH4 + concentration, and (d) the percentage of SO4 2- concentration compared to the initial dose in LB medium in response to different AMD doses from 0 to 2,500 mg SO4 2- /L. |
|
Fig. 2 Variation in (a) pH, (b) dehydrogenase activity, and (c) ammonium (NH4 +) concentration of soil slurry containing different amounts of acidic soil (i.e., 0, 1, 10, 25, 50, 60, 75, and 100 g) spiked by AMD for 6 days of incubation |
|
Fig. 3 Comparison of resulting (a) pH, (b) dehydrogenase
activity, and (c) NH4
+
concentration of soil slurry containing
different amount of sphagnum peat of 0, 0.1, 1, 2.5, and 5 g in an
acidic mine soil slurry for 6 days of incubation. |
|
Fig. 4 Correlation matrix among the variation of dehydrogenase activity, produced NH4 + and their resulting pH depending on the various soil contents of (a) acidic soil and (b) sphagnum peat, respectively |
|
Table 1 Two-way analysis of variance (ANOVA) for pH evolution in different experimental sets of varying amounts of acidic soil together with different sphagnum peat fraction according to incubation times |
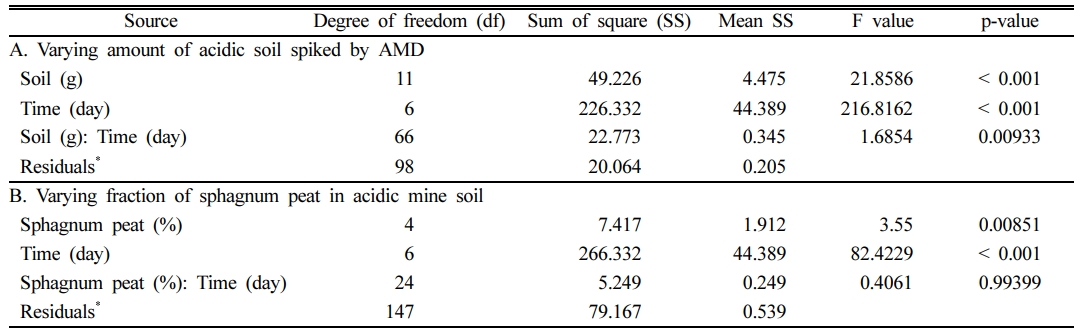
*. Residual is the difference between the observed and the estimated value in prediction model. |
This study highlights that acid-neutralizing bacterial consortium mainly composed of Klebsiella sp. and Raoultella sp. remediates severe AMD-impaired soil by effective improvement of the soil pH through microbial metabolism, in particular, ammonium production in the aqueous phase. However, high concentration of SO42- together with greater amount of humic substance (i.e., sphagnum peat) still have undermined the microbial activity under aerobic condition, leading to the less evolution of pH . In the meantime, the amount of acidic soil can be regarded as dominant factors in this microbe-mediated remediation process necessarily requiring for an increasing incubation times than the amount of sphagnum peat.
This work was supported by Korean Environment Industry & Technology Institute (KEITI) through Ecological Imitation-based Environmental Pollution Management Tech- nology Development Project, funded by Korea Ministry of Environment (MOE) (2019002800004) and partially supported by INHA University Grant.
- 1. Ahn, C.M. and Kim, C.G., 2015, Assessments of CO2 biomineralization and its kinetics using indigenous microorganisms derived from landfill cover soil, Desalin. Water Treat., 54, 3632-3638.
-
- 2. Alice, B., Melo, G. De, Motta, F.L., Helena, M., and Santana, A., 2016, Humic acids: structural properties and multiple functionalities for novel technological developments, Mater. Sci. Eng. C, 62, 967-974.
-
- 3. Asta, M.P., Ayora, C., Acero, P., and Cama, J., 2010, Field rates for natural attenuation of arsenic in Tinto Santa Rosa acid mine drainage (SW Spain), J. Hazard. Mater., 177, 1102-1111.
-
- 4. Barboza, N.R., Cordeiro, M.M., Santos, P.A., Queiroz, P.S., Leão, V.A., and Guerra-Sá, R., 2018, A new application for klebsiella oxytoca in bioremediation: treatment of manganese-laden wastewaters, Water. Air. Soil Pollut., 229, 19-26.
-
- 5. Boopathy, R., Gurgas, M., Ullian, J., and Manning, J.F., 1998, Metabolism of explosive compounds by sulfate-reducing bacteria, Curr. Microbiol., 37, 127-131.
-
- 6. Chemeda, Y.C., Deneele, D., and Ouvarard, G., 2018, Short-term lime solution-kaolinite interfacial chemistry and its effect on long-term pozzolanic activity, Appl. Clay Sci., 161, 419-426.
-
- 7. Costa, M.C. and Duarte, J.C., 2005, Bioremediation of acid mine drainage using acidic soil and organic wastes for promoting sulphate-reducing bacteria activity on a column reactor, Water. Air. Soil Pollut., 165, 325-345.
-
- 8. Fávere, V.T., Laus, R., Laranjeira, M.C.M., Martins, A.O., and Pedrosa, R.C., 2004, Use of chitosan microspheres as remedial material for acidity and iron (III) contents of coal mining wastewaters, Environ. Technol., 25, 861-866.
-
- 9. Freeman, C. and Lock, M.A., 1992, Recalcitrant high-molecular-weight material, an inhibitor of microbial metabolism in river biofilms, Appl. Environ. Microbiol., 58, 2030-2033.
-
- 10. Goulding, K.W.T., 2016, Soil acidification and the importance of liming agricultural soils with particular reference to the United Kingdom, Soil Use Manag., 32, 390-399.
-
- 11. Gupta, A. and Sar, P., 2020, Characterization and application of an anaerobic, iron and sulfate reducing bacterial culture in enhanced bioremediation of acid mine drainage impacted soil, J. Environ. Sci. Heal. - Part A Toxic/Hazardous Subst. Environ. Eng., 55, 464-482.
-
- 12. Haynes, R.J. and Mokolobate, M.S., 2001, Amelioration of Al toxicity and P deficiency in acid soils by additions of organic residues: a critical review of the phenomenon and the mechanisms involved, Nutr. Cycl. Agroecosystems, 59, 47-63.
-
- 13. Huang, Y.L., Yang, S., Long, G.X., Zhao, Z.K., Li, X.F., and Gu, M.H., 2016, Manganese toxicity in sugarcane plantlets grown on acidic soils of southern China, PLoS One, 11, 1-18.
-
- 14. Hubert, C. and Voordouw, G., 2007, Oil field souring control by nitrate-reducing Sulfurospirillum spp. that outcompete sulfate-reducing bacteria for organic electron donors, Appl. Environ. Microbiol., 73, 2644-2652.
-
- 15. Kefeni, K.K., Msagati, T.A.M., and Mamba, B.B., 2017, Acid mine drainage: Prevention, treatment options, and resource recovery: A review, J. Clean. Prod., 151, 475-493.
-
- 16. Lee, S.J., Park, J.H., Ahn, Y.T., and Chung, J.W., 2015, Comparison of heavy metal adsorption by peat moss and peat moss-derived biochar produced under different carbonization conditions, Water. Air. Soil Pollut., 226, 8-16.
-
- 17. Leiva, E., Leiva-Aravena, E., and Vargas, I., 2016, Acid water neutralization using microbial fuel cells: an alternative for acid mine drainage treatment, Water, 8, 536-544.
-
- 18. Liu, Z., Li, L., Li, Z., and Tian, X., 2018, Removal of sulfate and heavy metals by sulfate-reducing bacteria in an expanded granular sludge bed reactor, Environ. Technol. (United Kingdom), 39, 1814-1822.
-
- 19. Luptakova, A., Ubaldini, S., MacIngova, E., Fornari, P., and Giuliano, V., 2012, Application of physical-chemical and biological-chemical methods for heavy metals removal from acid mine drainage, Process Biochem., 47, 1633-1639.
-
- 20. Ma, J.F. and Furukawa, J., 2003, Recent progress in the research of external Al detoxification in higher plants: a minireview, J. Inorg. Biochem., 97, 46-51.
-
- 21. Marschner, P., Kandeler, E., and Marschner, B., 2003, Structure and function of the soil microbial community in a long-term fertilizer experiment, Soil Biol. Biochem., 35, 453-461.
-
- 22. Nancharaiah, Y.V., Mohan, T.V.K., Sai, P.M.S., and Venugopalan, V.P., 2017, Denitrification of high strength nitrate bearing acidic waters in granular sludge sequencing batch reactors, Inthernational Biodeterior. Biodegrad., 119, 29-36.
-
- 23. Núñez-Gómez, D., Rodrigues, C., Lapolli, F.R., and Lobo-Recio, M.Á., 2018, Adsorption of heavy metals from coal acid mine drainage by shrimp shell waste: isotherm and continuous-flow studies, J. Environ. Chem. Eng., 7, 102787-102796.
-
- 24. OECD (Organisation for Economic Co-operation and Development), 1984, Guidelines for testing of chemicals No. 207. Earthworm, acute toxicity test, OECD, Paris.
- 25. Ogbughalu, O.T., Gerson, A.R., Qian, G., Smart, R.S.C., Schumann, R.C., Kawashima, N., Fan, R., Li, J., and Short, M.D., 2017, Heterotrophic microbial stimulation through biosolids addition for enhanced acid mine drainage control, Minerals, 7, 105-116.
-
- 26. Okai, M., Suwa, C., Nagaoka, S., Obara, N., Mitsuya, D., Kurihara, A., Ishida, M., and Urano, N., 2017, Neutralization of acidic drainage by Cryptococcus sp. T1 immobilized in alginate beads, Biosci. Biotechnol. Biochem., 81, 2216-2224.
-
- 27. Olds, W.E., Tsang, D.C.W., Weber, P.A., and Weisener, C.G., 2013, Nickel and Zinc removal from acid mine drainage: roles of sludge surface area and neutralising agents, J. Min., 2013, 1-5.
-
- 28. Park, E.H., Mahanty, B., Yoon, S.U., and Kim, C.G., 2016, Biomimetic acid soil amelioration using indigenous soil isolates in comparison to other potential microorganism, Commun. Soil Sci. Plant Anal., 47, 1296-1305.
-
- 29. Qi, Y., Zhu, J., Fu, Q., Hu, H., and Huang, Q., 2017, Sorption of Cu by humic acid from the decomposition of rice straw in the absence and presence of clay minerals, J. Environ. Manage., 200, 304-311.
-
- 30. Qin, J., Cui, X., Yan, H., Lu, W., and Lin, C., 2019, Active treatment of acidic mine water to minimize environmental impacts in a densely populated downstream area, J. Clean. Prod., 210, 309-316.
-
- 31. Qiu, R., Zhao, B., Liu, J., Huang, X., Li, Q., Brewer, E., Wang, S., and Shi, N., 2009, Sulfate reduction and copper precipitation by a Citrobacter sp. isolated from a mining area, J. Hazard. Mater., 164, 1310-1315.
-
- 32. Rehr, B. and Klemme, J.H., 1989, Formate dependent nitrate and nitrite reduction to ammonia by Citrobacter freundii and competition with denitrifying bacteria, Antonie Van Leeuwenhoek, 56, 311-321.
-
- 33. Riaz, M., Yan, L., Wu, X., Hussain, S., Aziz, O., and Jiang, C., 2018, Mechanisms of organic acids and boron induced tolerance of aluminum toxicity: a review, Ecotoxicol. Environ. Saf., 165, 25-35.
-
- 34. Rodriguez-Navarro, C., Rodriguez-Gallego, M., Ben Chekroun, K., Gonzalez-Munoz, M.T., Chekroun, Koutar Ben, and Gonzalez-Muñoz, M.T., 2003, Conservation of ornamental stone by Myxococcus xanthus-induced carbonate biomineralization, Appl. Environ. Microbiol., 69, 2182-2193.
-
- 35. RoyChowdhury, A., Sarkar, D., and Datta, R., 2015, Remediation of acid mine drainage-impacted water, Curr. Pollut. Reports, 1, 131-141.
-
- 36. Sánchez-Andrea, I., Sanz, J.L., Bijmans, M.F.M., and Stams, A.J.M., 2014, Sulfate reduction at low pH to remediate acid mine drainage, J. Hazard. Mater., 269, 98-109.
-
- 37. Sasikala, C., Jiwal, S., Rout, P., and Ramya, M., 2012, Biodegradation of chlorpyrifos by bacterial consortium isolated from agriculture soil, World J. Microbiol. Biotechnol., 28, 1301-1308.
-
- 38. Sharma, S., Lee, M., Reinmann, C.S., Pumneo, J., Cutright, T.J., and Senko, J.M., 2020, Impact of acid mine drainage chemistry and microbiology on the development of efficient Fe removal activities, Chemosphere, 249, 126117.
-
- 39. Smilek, J., Sedláček, P., Kalina, M., and Klučáková, M., 2015, On the role of humic acids¡¯ carboxyl groups in the binding of charged organic compounds, Chemosphere, 138, 503-510.
-
- 40. Sugimori, D., Watanabe, M., and Utsue, T., 2013, Isolation and lipid degradation profile of Raoultella planticola strain 232-2 capable of efficiently catabolizing edible oils under acidic conditions, Appl. Microbiol. Biotechnol., 97, 871-880.
-
- 41. Tang, H., Pu, W.C., Cai, C.F., Xu, J.P., and He, W.J., 2016, Remediation of acid mine drainage based on a novel coupled membrane-free microbial fuel cell with permeable reactive barrier system, Polish J. Environ. Stud., 25, 107-112.
-
- 42. Tao, W., Kasuga, T., Li, S., Huang, H., and Fan, Z., 2019, Homoethanol production from cellobionate and glycerol using recombinant Klebsiella oxytoca strains, Biochem. Eng. J., 151, 107364.
-
- 43. Tejeda-agredano, M., Mayer, P., and Ortega-calvo, J., 2014, The effect of humic acids on biodegradation of polycyclic aromatic hydrocarbons depends on the exposure regime, Environ. Pollut., 184, 435-442.
-
- 44. Tikhonov, V.V., Yakushev, A.V., Zavgorodnyaya, Y.A., Byzov, B.A., and Demin, V.V., 2010, Effects of humic acids on the growth of bacteria, Eurasian Soil Sci., 43, 305-313.
-
- 45. Tolonen, E.T., Sarpola, A., Hu, T., Rämö, J., and Lassi, U., 2014, Acid mine drainage treatment using by-products from quicklime manufacturing as neutralization chemicals, Chemosphere, 117, 419-424.
-
- 46. Trevors, J.T., 1984, Effect of substrate concentration, inorganic nitrogen, O2 concentration, temperature and pH on dehydrogenase activity in soil, Plant Soil, 77, 285-293.
-
- 47. Vadapalli, V.R.K., Zvimba, J.N., Mathye, M., Fischer, H., and Bologo, L., 2015, Acid mine drainage neutralization in a pilot sequencing batch reactor using limestone from a paper and pulp industry, Environ. Technol., 36, 2515-2523.
-
- 48. Wang, D., Zheng, G., and Zhou, L., 2012, Isolation and characterization of a nitrobenzene-degrading bacterium Klebsiella ornithinolytica NB1 from aerobic granular sludge, Bioresour. Technol., 110, 91-96.
-
- 49. Yan, F. and Schubert, S., 1996, Soil pH increase due to biological decarboxylation of organic anions, Soil Biol. Biochem., 28, 617-624.
-
- 50. Yuan, J.H., Xu, R.K., Qian, W., and Wang, R.H., 2011, Comparison of the ameliorating effects on an acidic ultisol between four crop straws and their biochars, J. Soils Sediments, 11, 741-750.
-
- 51. Zhang, Xiaohui, Tang, S., Wang, M., Sun, W., Xie, Y., Peng, H., Zhong, A., Liu, H., Zhang, Xiaowei, Yu, H., Giesy, J.P., and Hecker, M., 2019, Acid mine drainage affects the diversity and metal resistance gene profile of sediment bacterial community along a river, Chemosphere, 217, 790-799.
-
 This Article
This Article
-
2021; 26(1): 34-44
Published on Feb 28, 2021
- 10.7857/JSGE.2021.26.1.034
- Received on Jan 6, 2021
- Revised on Jan 9, 2021
- Accepted on Feb 3, 2021
Services
- Abstract
1.introduction
2.materials and methods
3.results and discussion
4.conclusion
- Acknowledgements
- References
- Full Text PDF
Shared
Correspondence to
- Chang Gyun Kim
-
1Program in Environmental and Polymer Engineering, INHA University, Incheon 22212, Korea
2- E-mail: cgk@inha.ac.kr